Next: RNA folding and evolutionary
Up: research
Previous: Novelty in Evolution: Introduction
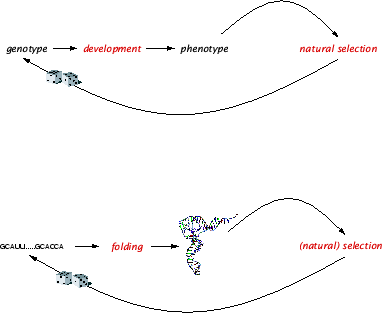
[1] The folding of RNA sequences into secondary structures. This is arguably the simplest, non-trivial, biophysically grounded instance of the novelty problem. The case is conceptually simple, because a formal characterization of both the possible genotypes (sequences) and phenotypes (shapes) is available along with a reasonably transparent model of the mapping between the two. This enables a systematic exploration of the variation of phenotype with genotype. The primary goal is not the most accurate possible prediction of the shape into which a particular sequence folds, but rather a catalog of statistical characteristics of the relationship between sequence and structure. In a wider context, one would like to know how these characteristics interact with evolution and whether they can be expected to hold at higher levels of phenotypic complexity.
Figure: The folding of RNA sequences into shapes as a proxy of a genotype-phenotype map. Mutations occur at the genetic level. Their consequences at the phenotypic level are mediated by development, the suite of processes by which phenotype is constructed from genotype. RNA folding is a transparent and tractable model that captures this indirection of innovation within a single molecule. The RNA folding map is characterized by a number of remarkable statistical regularities with profound evolutionary consequences. It is plausible that these regularities generalize to more complex forms of development.
 |
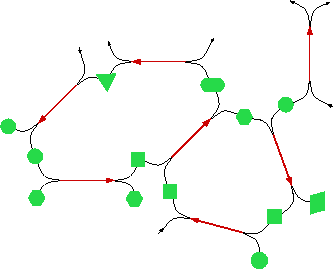
[2] A formal abstraction of chemistry and a characterization of the self-maintaining production networks it generates. Here I focus on networks of molecules that generate new molecules in chemical reactions. Leo Buss and I define as an ``organization'' a network that is self-maintaining. The organization of such a network consists in the specific functional relationships between molecules that collectively ensure their continuous regeneration. The problem here consists in formalizing chemical reactions between molecules or, more generally, interactions between agents that result in the construction of new agents.
Figure: Organization as a self-maintaining network of production pathways. Molecules (represented by green shapes) react (red arrows) to generate new molecules that react to generate further molecules. In this way, an initial collection of molecules generates a network of chemical transformations. A reaction can be viewed in analogy to a logical inference or derivation: the reactants are premises and the products are conclusions which can be premises for further inferences. Leo Buss and I call ``organizations'' those networks that have become self-maintaining under conditions in which each molecule has a finite lifetime (because of an outflow or degradation).
 |
This requires a theory linking the structure of agents to their behavior. In chemistry, the behavior of molecules consists in deterministically transforming the structure of those very molecules, thus yielding new behaviors. The feedback between agent structure which determines action and action targeted at agent structure generates, under appropriate boundary conditions, self-maintaining networks of production. The goal is to find and exploit a theory of such agents to characterize the networks they sustain. Ultimately, the question is: What is chemistry? Quantum mechanics answers that question by clarifying how chemistry arises from physics. Unfortunately, that answer does not clarify how molecular organization arises from chemistry. A different perspective on chemistry is needed, one that views molecules as grammatical entities encoding packets of action. Such a view must yield tools for plugging these actions together and for analyzing the resulting network closures under a variety of boundary conditions.
[3] Molecular signaling and the nature and behavior of distributed control networks. Biological organization is more than a self-maintaining closure of chemical transformations. Organization also means control, that is, starting, stopping, synchronizing and sequencing concurrent processes whose coordination underlies
Figure: Self-rewiring signaling networks. The upper portion depicts a network of signaling proteins that activate (green) or deactivate (red) one another. The communication targets of each protein are controlled by the activation state of other proteins in the same network. Changes in that activation state cause the wiring diagram to change. The lower portion depicts the receptor interface that functions as an input to the network. Upon binding specific signals, receptors become activated and propagate the signal into the network. The gene expression layer has been omitted.
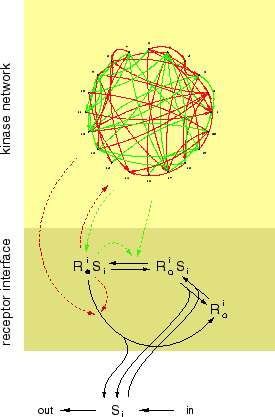 |
behavior. Control and coordination within and between cells is achieved by molecular communication networks that transduce and relay signals typically originating with the binding of molecules to receptors located in the cell membrane. Communication here means the activation or deactivation of a protein by another protein by transferring or removing a chemical token, such as a phosphate or methyl group. I want to explore molecular signal processing from a new angle by considering a network in which the communication links between components are not fixed, but controlled by the state of other components in the same network. This gives rise to a feedback between the network architecture and the activation kinetics induced on the network by that architecture. I further imagine a gene expression layer coupled to the network. By turning on and off genes that code for its own components, the network can control its own composition. The objective is to formulate a simple model system operating at three time scales with feedbacks among them: a short time scale at which the activation state of components changes, a medium time scale at which communication links change and a long time scale at which the very composition of the system is altered. I want to characterize the dynamical behavior of this class of systems, investigate their potential for learning and clarify the analogy to neural networks.
In the following I briefly elaborate on each project.
Subsections
Next: RNA folding and evolutionary Up: research Previous: Novelty in Evolution: Introduction
Walter Fontana, Santa Fe Institute
|
|
|